Introduction
Follicular lymphoma (FL) represents the most prevalent indolent B-cell lymphoma, comprising up to 25% of all non-Hodgkin lymphoma cases [1]. A combination of anti-CD20 agents with chemotherapy in the frontline setting results in favorable initial treatment responses in the majority of patients [2-6], leading to overall survival rates rivaling normal life expectancy [7]. Importantly, a significant number of patients follow clinical trajectories that are distinct from the otherwise prolonged and indolent course. In particular, approximately 20% experience early disease progression (commonly referred to as progression of disease within 24 months; POD24), which is associated with markedly inferior clinical outcomes [8-11]. A sizeable minority also undergoes histological transformation into an aggressive lymphoma, most commonly diffuse large B-cell lymphoma (DLBCL), a complication likewise linked to rapid progression and inferior survival [12-14].
Technological breakthroughs in the fields of genomics, transcriptomics, and epigenomics have advanced the understanding of FL from being a single entity primarily driven by the t(14;18) translocation to a highly heterogeneous group of tumors with diverse molecular features [15]. These advances have also provided a rationale for the introduction of multiple novel therapeutic agents which target the various pathways related to the underlying disease pathogenesis, including epigenetic regulators [16, 17], the B-cell receptor (BCR) pathway [18, 19], E3 ubiquitin ligase complexes [20, 21] and the tumor microenvironment (TME) - via T-cell engagers such as bispecific antibodies [22-25], chimeric antigen receptor (CAR) T cells [26-28], or via promoting tumor cell phagocytosis [29].
In view of the expanding range of novel treatment options, it is essential to reliably identify the high-risk patients who are unlikely to benefit from conventional chemoimmunotherapy, such as those predisposed to early progression or transformation. Although a number of clinical predictors were shown to be associated with these poor outcomes, including advanced stage, elevated LDH, poor performance status, presence of B symptoms, extranodal site involvement, and high overall Follicular Lymphoma International Prognostic Index (FLIPI) score [9, 30-33], none of these risk factors have sufficient pre-treatment predictive capacity. Moreover, the biologic events that shape a particular disease trajectory in FL, and, therefore, have the potential to serve as candidate biomarkers for poor clinical outcomes, are still incompletely understood. Although genomic studies have provided a foundation for the mechanisms of FL lymphomagenesis, it has become increasingly evident that the disease cannot be fully explained at the DNA level alone, highlighting the need to investigate additional layers of tumor biology. These challenges are currently being addressed through the use of novel techniques such as single-cell profiling and multiomics, which allow for a more comprehensive interrogation of both the malignant and non-malignant tumor compartments. In this review, a brief clinician-oriented overview is provided of how these and other techniques are reshaping our understanding of de novo and transformed FL, presents a rationale for how this knowledge may eventually translate into clinically meaningful risk assessment and therapy development.
Materials and methods
The PubMed/MEDLINE database was searched using combinations of terms including “follicular lymphoma”, “transformation”, “diffuse large B-cell lymphoma”, “POD24”, “early progression”, “tumor microenvironment”, “single-cell”, “spatial”, “integrated profiling” and “multiomics.” Reference lists of publications were manually searched to identify additional relevant studies. Titles and abstracts were screened for relevance, followed by full-text review of potentially eligible articles. This review considered peer-reviewed original research, reviews and conference abstracts addressing FL and its histological transformation to DLBCL. Non–peer-reviewed sources and isolated case reports were excluded.
Results
The role of genomic sequencing in follicular lymphoma
BCL2 dysregulation. Before the advent of next-generation sequencing (NGS) technologies, the knowledge of specific events leading to FL development was largely limited to a single genetic lesion (Fig. 1). Early cytogenetic studies from the mid-1980s revealed that 85-90% of FL patients carry a hallmark reciprocal translocation involving the B-cell lymphoma 2 (BCL2) oncogene on chromosome 18 and the immunoglobulin heavy chain (IgH) gene locus on chromosome 14 [34-36]. As a consequence, the BCL2 gene is placed under the control of strong IgH gene enhancer elements, resulting in constitutive BCL2 overexpression [37]. The BCL2 gene product then acts as its role as an anti-apoptotic protein, providing a selective survival advantage to the FL precursors carrying the t(14;18) translocation [38] and allowing them to accumulate secondary genetic lesions without undergoing apoptosis [39].
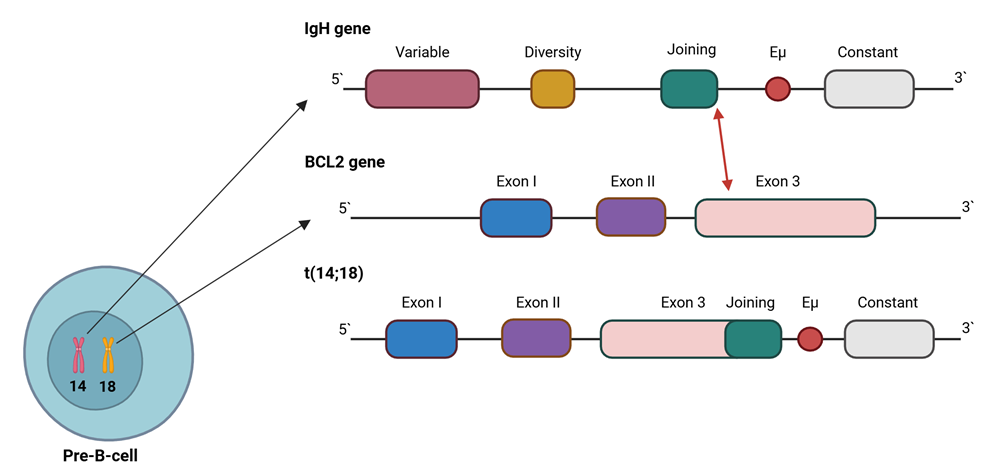 |
Fig. 1 A simplified schematic depiction of the hallmark t(14;18) rearrangement. Note: The reciprocal translocation, arising during the physiological V(D)J recombination process in developing B cells, juxtaposes BCL2 (chromosome 18, typically involving the region adjacent to exon 3) to the IgH (chromosome 14) Joining (J) region (red arrow), resulting in constitutive BCL2 expression driven by IgH regulatory sequences (such as Eµ) and promoting apoptosis resistance in B cells carrying the rearrangement. |
More recently, a role for somatic mutations in the BCL2 gene has also been demonstrated. Correia et al. used Sanger sequencing to characterize BCL2 mutations in FL and reported that such mutations, identified in 12% of patients, represent an independent risk factor for transformation and death from lymphoma [40]. However, the reliance on conventional Sanger sequencing likely underestimated the true prevalence and spectrum of BCL2 alterations. Because Sanger sequencing produces only a single continuous read per DNA fragment, it is not sensitive enough to reliably detect mutations present below a variant allele frequency (VAF) of 15-20% [41]. This limitation is particularly important in the context of heterogeneous tumor samples, where a high background of DNA coming from normal cells within the tumor could mask mutations of the malignant clone. In contrast, NGS allows detection of less common variants through redundant sequencing of the same genomic regions. In line with the above, in a subsequent study by the same group employing NGS, BCL2 mutations were found in 52% of newly diagnosed FL [42], in stark contrast to the 12% prevalence reported using Sanger sequencing [40]. Beyond sensitivity, Sanger sequencing approaches are also inherently targeted, focusing on preselected genes or regions, and therefore may miss pathogenic alterations outside the interrogated targets. Importantly, the observation that the t(14;18) translocation can be detected in a fraction of B cells in the majority of healthy individuals [43] further supports the notion that BCL2 deregulation alone is insufficient for malignant transformation, and that additional oncogenic events are required for FL development.
Epigenetic modifiers. Early NGS studies were foundational in defining the recurrent mutational landscape of FL. In a seminal study, Morin et al. conducted whole genome sequencing (WGS) of FL and DLBCL tumor samples, identifying recurrent mutations in EZH2, a gene encoding a H3K27 histone methyltransferase [44]. In a subsequent study by the same group, the authors discovered frequent inactivating mutations in the KMT2D (MLL2) gene, encoding for another histone methyltransferase [45], in 89% of FL patients [46]. In parallel, Pasqualucci et al. reported frequent mutations in the functionally related histone/protein lysine acetyltransferases CREBBP and EP300, detected in 32.6% and 8.7% of FL samples, respectively [47]. Building on these observations, Okosun et al. conducted WGS or whole exome sequencing (WES) of multiple samples that were collected from 10 patients at different time points during the course of their disease [48]. This sequential approach enabled the construction of phylogenetic trees, reflecting the clonal evolution of FL for each patient. The analysis demonstrated a branching evolution pattern in each of the trees, with the “trunk” representing the putative driver lesions that are shared between the initially identified and the subsequent clones in the FL tumors, thus supporting the existence of a common precursor and divergent subclones. In all patients, the precursor populations demonstrated an enrichment for mutations in the above-mentioned epigenetic modifiers, including KMT2D, CREBBP, EP300, and EZH2, and these findings were validated in an additional cohort of over 100 FL biopsies, where concurrent mutations in at least 2 epigenetic modifiers were found in more than 70% of cases. Interestingly, a small number of patients showed distinct mutations between diagnostic and subsequent biopsies that occurred within the same genes, a finding that was recently also observed in DLBCL [49], suggesting that certain driver lesions may be indispensable for lymphomagenesis and determine the commitment of the tumor towards a particular genetic phenotype.
Genetic determinants of FL transformation to DLBCL. The above data established epigenetic dysregulation as a central driving mechanism in the pathogenesis of de novo FL, with oncogenic effects that include immune evasion, cell cycle dysregulation, and alteration of multiple signaling pathways, including BCR, JAK-STAT, and NF-κB [15]. Importantly, analysis of sequential patient biopsies in the study of Okosun et al. reported that other genetic events, particularly abnormalities affecting cell cycle regulation and apoptosis (e.g., MDM2, MYC), as well as NF-κB signaling (REL, MYD88, TNFAIP3), appeared by the time of histological transformation of FL into DLBCL, and were not detectable in the initial FL biopsies [48]. Consistent with this, subsequent studies have demonstrated an association between transformation and increased mutational burden, in addition to recurrent genetic lesions affecting immune escape (B2M), confinement of B cells to the germinal center (GNA13, S1PR2, P2RY8), and cell cycle regulation and apoptosis (CDKN2A, CDKN2B, TP53,MYC) [50-53]. Notably, such abnormalities, in particular those affecting CDKN2A/2B, TP53 and MYC have also been implicated in transformation of chronic lymphocytic leukemia into DLBCL [54-56], and represent defining features of several prognostically inferior genetic subtypes of de novo DLBCL [53, 57-59]. Collectively, these findings support the notion that acquisition of specific genetic events contributes to the transformation of FL (Fig. 2) and raise the possibility that similar mechanisms play an important role in the development of more aggressive phenotypes across a spectrum of lymphoid malignancies [60].
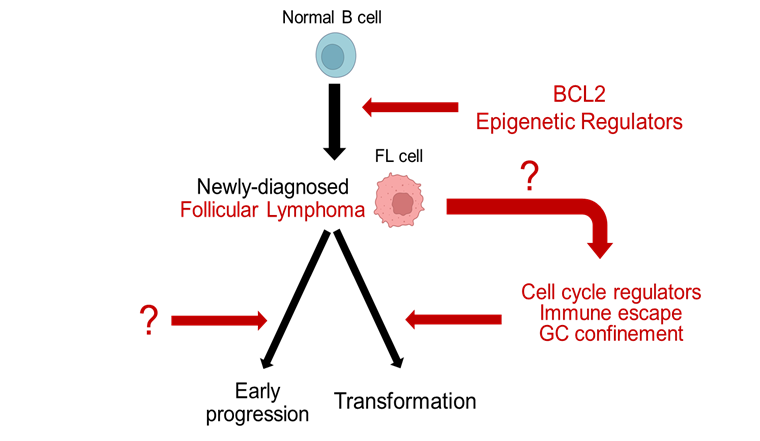 |
Fig. 2 Conceptual model of follicular lymphoma pathogenesis. Note: Early molecular events, including BCL2 deregulation and recurrent alterations in epigenetic regulators (upper right), contribute to malignant transformation of normal B cells into follicular lymphoma. Following diagnosis, FL may follow distinct evolutionary and clinical courses, including high-risk outcomes such as early progression and histological transformation to aggressive lymphoma. Transformation is accompanied by gain of additional genetic lesions, such as those affecting cell cycle regulators, immune escape, and germinal center confinement (lower right). Black arrows indicate established evolutionary relationships; red arrows indicate the mechanisms driving these relationships. Question marks denote biologic events that are incompletely understood: the determinants of early progression (lower left) and the events leading to acquisition of transformation-enabling genetic lesions (middle right). |
The above findings indicate that transformation of FL into DLBCL is driven by an expansion of a tumor subclone with the propensity to acquire transformation-enabling genetic lesions [52, 61], a tendency that may arise as a functional consequence of pre-existing genetic abnormalities or other biologic events. In this context, it appears to be of more value to identify how the mutations at initial diagnosis influence the risk of FL transformation, rather than those acquired later during the course of disease progression, given that such an approach could eventually help guide up-front clinical decision-making. In a recent study that analyzed WGS data from 423 FL and de novo DLBCL patients, two genetically distinct FL subgroups were resolved using a machine learning classifier trained to discriminate between DLBCL and FL based on mutational profiles [62]. The first group, which included 53% of untransformed FL from the training cohort, was termed the constrained FL (cFL). Patients within this group were less likely to undergo transformation, and were distinguished by the presence of missense mutations in the lysine acetyltransferase domain of CREBBP, in addition to mutations in several genes involved in the mTORC1 signaling pathway (RRAGC, ATP6AP1, and ATP6V1B2). The remaining 47% of untransformed FL were classified as DLBCL-like FL (dFL), a subgroup presenting with increased rates of aberrant somatic hypermutation (as demonstrated by a higher prevalence of mutations within the transcription start sites of commonly affected genes, such as BCL6, BCL7A, RHOH, and ZFP36L1), nonsense CREBBP mutations, and higher risk of transformation to DLBCL. These findings indicate that certain early genetic lesions may predispose FL to a commitment toward histological transformation, and thus serve as potential predictors for this clinical outcome. Whether these early genetic events contribute to transformation directly, or determine a state in which additional transformation-inducing genetic events accumulate, is currently unclear.
Genetic risk stratification models. Data from the aforementioned studies have laid the foundation for developing pretreatment prognostic scoring systems that incorporate somatic mutations for FL risk stratification. The most well-known of these models is the m7-FLIPI, a molecular prognostic tool created using a targeted NGS panel of 74 genes applied to pretreatment biopsies from 151 FL patients [63]. The score integrates several clinical parameters with the mutation status of 7 genes: EZH2, ARID1A, MEF2B, EP300, FOXO1, CREBBP, and CARD11. In the initial trials, it was able to stratify patients into low- and high-risk subgroups based on failure-free survival [63], as well as identify those at highest risk for POD24 [9]. However, multiple follow-up studies attempting to validate the m7-FLIPI score have yielded inconsistent results in external patient cohorts [33, 64-67], suggesting that specific genetic lesions may be associated with distinct functional consequences in different clinical and biologic contexts. Given these challenges, no mutation-based prognostic score is currently adopted in clinical practice. As expanded upon below, the extensive inter- and intraclonal heterogeneity inherent to FL likely diminishes the ability of simplified models that are based on individual genes to consistently identify high-risk patients across different settings. At earlier time points, such as at diagnosis, the smaller, potentially prognostically relevant subclones may, at best, only be detectable using ultra-sensitive technologies [52, 61]. Moreover, none of the existing risk-stratification models were specifically trained for the most pertinent predictor of inferior survival, POD24 [68]. Finally, the risk of adverse clinical outcomes, such as transformation, may be associated with exposure to specific treatment regimens [69], further complicating our understanding of the predictive value of genetic lesions in FL. Collectively, these findings indicate that conventional, bulk DNA-level profiling alone is not sufficient to identify the core determinants underlying the clinical trajectories seen in FL.
The role of single-cell sequencing and integrated multiomic approaches in follicular lymphoma
Single-cell profiling and tumor microenvironment as a driver of clinical behavior. Bulk DNA sequencing is readily accessible, and widely used across cancer studies for diagnosis and biomarker identification. However, every tumor cell can exhibit a unique genomic, epigenomic, and transcriptional profile. Modifications at the RNA level appear particularly important, given that genomic and epigenomic changes are ultimately reflected at the RNA level. With the development of the 10x Genomics Chromium systems, single-cell RNA sequencing has recently become more accessible to the scientific community, allowing for analysis of gene expression profiles from thousands of individual cells per run [70], exposing subclones and cell states that would otherwise be invisible in bulk sequencing.
Multiple studies employing single-cell RNA sequencing have reaffirmed that FL tumors consist of several coexisting subclones, either at different anatomical sites, or within the same lesion. In a recent study, Haebe et al. used single-cell RNA, B- and T-cell receptor sequencing, as well as flow cytometry to profile synchronously acquired tumors from different sites in 10 patients with FL [71]. Unexpectedly, the authors found that in many patients the disease evolved independently at different sites, exhibiting site-to-site divergence in BCR evolution, gene expression and surface protein profiles. Supporting these observations, bulk WES of samples obtained from multiple disease sites within the same patients revealed spatially discordant genetic abnormalities, including alterations in several m7-FLIPI-related genes, such as EZH2 and EP300 [72]. These findings appear particularly relevant given that most FL patients manifest disseminated tumor involvement. They also support the notion that a diagnostic biopsy from a single anatomical site may be insufficient, and further add to the limited predictive ability of the approaches relying just on bulk genome sequencing and clinical data. Moving forward, the potential requirement for multiple biopsies may be circumvented by genomic profiling of circulating tumor DNA [73], although the utility of such an approach in FL remains to be validated.
In a study integrating single-cell RNA and bulk exome sequencing, Andor et al. also identified the presence of multiple subclones within individual FL tumors, each associated with transcriptional profiles that reflected their distinct genetic backgrounds [74]. Notably, substantial transcriptional heterogeneity was also observed within individual genetic subclones, suggesting that phenotypic diversity is not solely determined by the genotype. Consistent with this, malignant B cells in FL were shown to cycle between different transcriptional states largely independently of their mutational profile, a process likely driven by extrinsic signals from the non-malignant components of the lymphoma microenvironment [75].
The structure of lymph nodes affected by FL retains features of normal lymphoid tissue, but the architecture is disorganized, with complete or partial effacement by the neoplastic follicles [76]. It is enriched for T cells, including mostly CD4+ subsets, such as T follicular helper cells, T regulatory cells, T follicular regulatory cells, and others, which were shown to contribute to tumor growth and treatment resistance via pro-survival signals (such as CD40L or IL-4), or by suppressing the normal anti-tumor immune response. Other cell types include the various macrophages and dendritic cells, which are frequently polarized into a tumor-supportive phenotype by the malignant cells, as well as a variety of stromal cells, which are able to promote tumor growth by altering the microenvironment or by interacting directly with the tumor cells and secreting tumor-promoting cytokines. Dave et al. were among the first to demonstrate the role of the FL TME in determining the patient clinical trajectory [77]. Using microarray-based gene-expression profiling, the authors resolved two survival-associated gene expression signatures, each reflecting the biologic characteristics of the non-malignant cells within the analyzed tumors. The first, termed immune-response 1, was associated with a favorable prognosis and enriched for expression of genes encoding T-cell and macrophage markers, whereas the second, termed immune-response 2, was associated with a poor prognosis and enriched only for genes preferentially expressed in macrophages and dendritic cells, thus suggesting a role for the TME-derived T cells in preventing tumor growth. These early bulk gene-expression data provided the first evidence that the FL microenvironment is a prognostically meaningful structure that could shape the disease trajectory. Single-cell technologies have since expanded this concept substantially. Han et al. used single-cell RNA sequencing to subdivide FL into four major subtypes based on the phenotype and relative abundance of the various T cell populations [78]. Importantly, the subtype characterized by T cell depletion was associated with poor survival, reinforcing the role of T cells in suppressing tumor growth and suggesting that immune evasion by the tumor cells is an important contributor to poor prognosis.
Integrated multiomic approaches in early relapse FL and transformation to DLBCL. A multiomic approach refers to the integrated analysis of two or more biologic data layers (e.g., genomics, transcriptomics, epigenomics, proteomics) to reconstruct a more complete view of tumor biology. In FL, where clonal heterogeneity and microenvironmental interactions appear to significantly influence disease behavior, multiomic profiling is particularly valuable for capturing the complexity that may otherwise be missed by single-modality methods. Using WES, bulk and single-cell RNA sequencing, and iterative bleaching extends multiplexity (IBEX) imaging (an immunofluorescence technique that allows visualization of more than 65 proteins in the same tissue section) [79], Radtke et al. constructed a molecular and cellular atlas of lymph nodes affected by FL [80]. The authors demonstrated that malignant B cells in high-risk patients undergoing early relapse exhibited increased expression of genes related to BCR signaling and TME remodeling, unlike the malignant B cells derived from all other FL patients. The unique imaging technique enabled spatial resolution of the non-malignant cell populations within individual tumors, revealing that early-relapse cases were characterized by an expansion of specific stromal cell communities and desmoplasia. Notably, the tumor B cells were frequently found to be in close physical proximity to certain TME components (such as dendritic cells), a state that may facilitate sustained BCR signaling.
Additional insights were obtained from another integrative single-cell study by Sarkozy et al., who applied single-cell genome and transcriptome sequencing to investigate the co-evolution of the malignant B cells and the surrounding microenvironment during transformation of FL into DLBCL [61]. Transcriptomic profiling showed differential expression of MYC target genes, in addition to activation of oxidative phosphorylation and mTORC1 pathways during transformation. By integrating WGS and single-cell RNA sequencing data across sequential time points, the authors further identified an association between genomic evolution (i.e., emergence of additional genetic lesions) and acquisition of a transformation-related transcriptional phenotype. However, significant transcriptional diversity in the absence of detectable genomic evolution was also observed, reinforcing the notion that genotype alone does not fully determine the tumor cell state. Analysis of the non-B cell fraction revealed marked remodeling of the TME, with decreases in naive/memory-like and follicular helper-like T cells, accompanied by expansion of the exhausted/regulatory-like and effector T-cell clusters, findings that were further supported by immunofluorescence evidence of increased CD8+ T-cell exhaustion marker expression during transformation. Spatial profiling additionally showed that the exhausted T cells were more abundant in close proximity to the malignant B cells after transformation, suggesting that the malignant B cells engaging with the surrounding T cells may contribute to T-cell dysfunction, and, potentially, to transformation-associated transcriptional reprogramming in the tumor B cells. Importantly, expression of the exhaustion marker LAG3 on CD8+ T cells in two independent pre-treatment FL cohorts was significantly associated with shorter time to transformation, indicating that specific TME features could represent candidate biomarkers for histological transformation. Collectively, these studies support the notion that both transformation and early relapse are accompanied by a shift in the composition and spatial organization of the TME, thus indicating that the tumor-microenvironment crosstalk is a core component of the biologic process that drives the development of more aggressive disease phenotypes (Fig. 3).
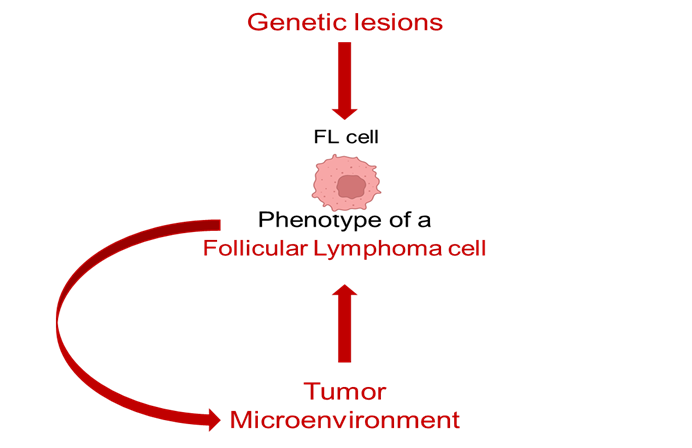 |
Fig. 3 Conceptual model for the determinants of a follicular lymphoma cell phenotype. Note: The phenotype of a malignant follicular lymphoma cell is shaped by both intrinsic genetic lesions (top arrow) and extrinsic signals from the tumor microenvironment (bottom arrow). The curved arrow denotes bidirectional crosstalk between tumor cells and surrounding non-malignant components, emphasizing the dynamic and reciprocal process determining the tumor cell phenotype. |
Discussion
Over the last few decades, the application of omics technologies has transformed the understanding of follicular lymphoma from a disease centered on BCL2 deregulation to a complex and heterogeneous entity shaped by multiple molecular mechanisms. Genomic sequencing has established epigenetic dysregulation as a core component of FL pathogenesis, with recurrent mutations in chromatin modifiers like KMT2D, CREBBP, EP300 and EZH2 occurring early and being maintained throughout disease evolution. In contrast, other genetic lesions, including those affecting cell cycle regulators, are more often acquired later on and are associated with more aggressive disease phenotypes such as transformation into DLBCL.
More recently, integrative approaches that combine genomics, transcriptomics, and spatial profiling have further shown that the mechanisms driving FL are multifaceted and cannot be explained by the genotype alone, instead reflecting the interplay between genetic lesions, transcriptional programs, and dynamic crosstalk with the lymphoma microenvironment. Collectively, the data presented in this review support a model of FL pathogenesis in which malignant cells undergo genetically driven phenotypic changes and engage in dynamic, bidirectional interactions with surrounding immune cells, thereby remodeling the immune microenvironment into a tumor-supporting framework that promotes further phenotypic changes in the malignant cells.
Despite these advances, FL continues to pose significant clinical challenges, with markedly inferior outcomes in patients with histological transformation or early progression. Recent discoveries, including the identification of LAG3+CD8+ T-cell infiltration as a candidate biomarker of histological transformation and the emergence of potential therapeutic vulnerabilities in early relapse, such as fibrosis-associated TME remodeling that may be targetable with antifibrotic agents and BCR pathway inhibitors, support the notion that multiomic profiling may continue to reveal clinically actionable biomarkers of high-risk disease and inform risk-adapted therapeutic strategies.
Conclusions
Follicular lymphoma is a biologically complex disease in which clinical outcomes are determined not only by the genetic background of the malignant cells, but also by their transcriptional states and interactions with the surrounding microenvironment. The data reviewed here indicate that continued research focused on multiomic profiling is essential to identify the distinct biologic determinants that drive the various clinical trajectories in FL, thereby improving our ability to predict poor outcomes and facilitating therapeutic approaches that are focused on timely prevention of transformation and early progression.
Competing interests
None declared.
Authors’ contributions
IN, OA and SB participated in conceptualization of the manuscript. IN conducted the literature review and drafted the manuscript. OA and SB critically revised the manuscript. All authors approved the final version of the manuscript.
Ethics approval
Not needed for this study.
Provenance and peer review
Not commissioned, externally peer-reviewed.
Declaration of Generative AI and AI-assisted technologies in the writing process
The GPT-5.2 model was used during copy-editing of the finished manuscript. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of this publication.
Authors’ ORCID IDs
Ivan Negara – https://orcid.org/0000-0002-1901-5187
Oleg Arnaut – https://orcid.org/0000-0002-5483-8672
Sanda Buruiana – https://orcid.org/0000-0003-2341-0099
References
Cerhan JR. Epidemiology of follicular lymphoma. Hematol Oncol Clin North Am. 2020 Aug;34(4):631-46. doi: 10.1016/j.hoc.2020.02.001.
Marcus R, Davies A, Ando K, Klapper W, Opat S, Owen C, et al. Obinutuzumab for the first-line treatment of follicular lymphoma. N Engl J Med. 2017 Oct 5;377(14):1331-44. doi: 10.1056/NEJMoa1614598.
Bachy E, Seymour JF, Feugier P, Offner F, López-Guillermo A, Belada D, et al. Sustained progression-free survival benefit of rituximab maintenance in patients with follicular lymphoma: long-term results of the PRIMA study. J Clin Oncol. 2019 Nov;37(31):2815-24. doi: 10.1200/JCO.19.01073.
Townsend W, Hiddemann W, Buske C, Cartron G, Cunningham D, Dyer MJS, et al. Obinutuzumab versus rituximab immunochemotherapy in previously untreated iNHL: final results from the GALLIUM study. HemaSphere. 2023 Jul;7(7):e919. doi: 10.1097/HS9.0000000000000919.
Kahl BS, Jegede OA, Peterson C, Swinnen LJ, Habermann TM, Schuster SJ, et al. Long-term follow-up of the RESORT study (E4402): a randomized phase III comparison of two different rituximab dosing strategies for low-tumor burden follicular lymphoma. J Clin Oncol. 2024 Mar 1;42(7):774-8. doi: 10.1200/JCO.23.01912.
Schulz H, Bohlius J, Skoetz N, Trelle S, Kober T, Reiser M, et al. Chemotherapy plus rituximab versus chemotherapy alone for B-cell non-Hodgkin’s lymphoma. Cochrane Database Syst Rev. 2007 Oct 17;2007(4):CD003805. doi: 10.1002/14651858.CD003805.pub2.
Tan D, Horning SJ, Hoppe RT, Levy R, Rosenberg SA, Sigal BM, et al. Improvements in observed and relative survival in follicular grade 1-2 lymphoma during 4 decades: the Stanford University experience. Blood. 2013 Aug 8;122(6):981-7. doi: 10.1182/blood-2013-03-491514.
Casulo C, Byrtek M, Dawson KL, Zhou X, Farber CM, Flowers CR, et al. Early relapse of follicular lymphoma after rituximab plus cyclophosphamide, doxorubicin, vincristine, and prednisone defines patients at high risk for death: an analysis from the national lymphocare study. J Clin Oncol. 2015 Aug 10;33(23):2516-22. doi: 10.1200/JCO.2014.59.7534.
Jurinovic V, Kridel R, Staiger AM, Szczepanowski M, Horn H, Dreyling MH, et al. Clinicogenetic risk models predict early progression of follicular lymphoma after first-line immunochemotherapy. Blood. 2016 Aug 25;128(8):1112-20. doi: 10.1182/blood-2016-05-717355.
Maurer MJ, Bachy E, Ghesquières H, Ansell SM, Nowakowski GS, Thompson CA, et al. Early event status informs subsequent outcome in newly diagnosed follicular lymphoma. Am J Hematol. 2016 Nov;91(11):1096-101. doi: 10.1002/ajh.24492.
Casulo C, Dixon JG, Le-Rademacher J, Hoster E, Hochster HS, Hiddemann W, et al. Validation of POD24 as a robust early clinical end point of poor survival in FL from 5225 patients on 13 clinical trials. Blood. 2022 Mar 17;139(11):1684-93. doi: 10.1182/blood.2020010263.
Hori Y, Hosoi H, Hiroi T, Wan K, Murata S, Morimoto M, et al. Utilizing clinical transformation criteria for prognostic stratification in follicular lymphoma prior to initial immunochemotherapy. Hematol Rep. 2024 Oct 4;16(4):612-23. doi: 10.3390/hematolrep16040060.
Mozessohn L, Cheung MC, Crump M, Buckstein R, Berinstein N, Imrie K, et al. Chemoimmunotherapy resistant follicular lymphoma: predictors of resistance, association with transformation and prognosis. Leuk Lymphoma. 2014 Nov;55(11):2502-7. doi: 10.3109/10428194.2014.885513.
Sarkozy C, Trneny M, Xerri L, Wickham N, Feugier P, Leppa S, et al. Risk factors and outcomes for patients with follicular lymphoma who had histologic transformation after response to first-line immunochemotherapy in the PRIMA trial. J Clin Oncol. 2016 Aug 1;34(22):2575-82. doi: 10.1200/JCO.2015.65.7163.
Araujo-Ayala F, Béguelin W. Biology as vulnerability in follicular lymphoma: genetics, epigenetics, and immunogenetics. Blood. 2025 Oct 9;146(15):1759-69. doi: 10.1182/blood.2024026020.
Morschhauser F, Tilly H, Chaidos A, McKay P, Phillips T, Assouline S, et al. Tazemetostat for patients with relapsed or refractory follicular lymphoma: an open-label, single-arm, multicentre, phase 2 trial. Lancet Oncol. 2020 Nov;21(11):1433-42. doi: 10.1016/S1470-2045(20)30441-1.
Shi Y, Gui L, Cheng Y, Wang H, Cao J, Li Y, et al. A phase II multicenter study of abexinostat, an oral histone deacetylase inhibitor, in patients with relapsed/refractory follicular lymphoma. J Clin Oncol. 2024 Jun;42(16_suppl):7059-7059. doi: 10.1200/JCO.2024.42.16_suppl.7059.
Zinzani PL, Mayer J, Flowers CR, Bijou F, De Oliveira AC, Song Y, et al. ROSEWOOD: a phase II randomized study of zanubrutinib plus obinutuzumab versus obinutuzumab monotherapy in patients with relapsed or refractory follicular lymphoma. J Clin Oncol. 2023 Nov 20;41(33):5107-17. doi: 10.1200/JCO.23.00775.
Shah NN, Zinzani PL, Wang ML, Nasta SD, Lech-Maranda E, Ogawa Y, et al. Pirtobrutinib, a highly selective, non-Ccvalent (reversible) BTK inhibitor in relapsed/refractory follicular lymphoma: results from the phase 1/2 BRUIN study. Blood. 2023 Nov 2;142(Suppl 1):3026. https://doi.org/10.1182/blood-2023-181309
Leonard JP, Trneny M, Izutsu K, Fowler NH, Hong X, Zhu J, et al. AUGMENT: a phase III study of lenalidomide plus rituximab versus placebo plus rituximab in relapsed or refractory indolent lymphoma. J Clin Oncol. 2019 May 10;37(14):1188-99. doi: 10.1200/JCO.19.00010.
Morschhauser F, Fowler NH, Feugier P, Bouabdallah R, Tilly H, Palomba ML, et al. Rituximab plus lenalidomide in advanced untreated follicular lymphoma. N Engl J Med. 2018 Sep 6;379(10):934-47. doi: 10.1056/NEJMoa1805104.
Linton KM, Vitolo U, Jurczak W, Lugtenburg PJ, Gyan E, Sureda A, et al. Epcoritamab monotherapy in patients with relapsed or refractory follicular lymphoma (EPCORE NHL-1): a phase 2 cohort of a single-arm, multicentre study. Lancet Haematol. 2024 Aug;11(8):e593-605. doi: 10.1016/S2352-3026(24)00166-2.
Budde LE, Sehn LH, Matasar M, Schuster SJ, Assouline S, Giri P, et al. Safety and efficacy of mosunetuzumab, a bispecific antibody, in patients with relapsed or refractory follicular lymphoma: a single-arm, multicentre, phase 2 study. Lancet Oncol. 2022 Aug;23(8):1055-65. doi: 10.1016/S1470-2045(22)00335-7.
Hutchings M, Morschhauser F, Iacoboni G, Carlo-Stella C, Offner FC, Sureda A, et al. Glofitamab, a novel, bivalent CD20-targeting T-cell-engaging bispecific antibody, induces durable complete remissions in relapsed or refractory B-cell lymphoma: a phase I trial. J Clin Oncol. 2021 Jun 20;39(18):1959-70. doi: 10.1200/JCO.20.03175.
Bannerji R, Arnason JE, Advani RH, Brown JR, Allan JN, Ansell SM, et al. Odronextamab, a human CD20×CD3 bispecific antibody in patients with CD20-positive B-cell malignancies (ELM-1): results from the relapsed or refractory non-Hodgkin lymphoma cohort in a single-arm, multicentre, phase 1 trial. Lancet Haematol. 2022 May;9(5):e327-39. doi: 10.1016/S2352-3026(22)00072-2.
Neelapu SS, Chavez JC, Sehgal AR, Epperla N, Ulrickson M, Bachy E, et al. Three-year follow-up analysis of axicabtagene ciloleucel in relapsed/refractory indolent non-Hodgkin lymphoma (ZUMA-5). Blood. 2024 Feb 8;143(6):496-506. doi: 10.1182/blood.2023021243.
Fowler NH, Dickinson M, Dreyling M, Martinez-Lopez J, Kolstad A, Butler J, et al. Tisagenlecleucel in adult relapsed or refractory follicular lymphoma: the phase 2 ELARA trial. Nat Med. 2022 Feb;28(2):325-32. doi: 10.1038/s41591-021-01622-0.
Morschhauser F, Dahiya S, Palomba ML, Martin Garcia-Sancho A, Reguera Ortega JL, Kuruvilla J, et al. Lisocabtagene maraleucel in follicular lymphoma: the phase 2 TRANSCEND FL study. Nat Med. 2024 Aug;30(8):2199-207. doi: 10.1038/s41591-024-02986-9.
Mehta A, Popplewell L, Collins GP, Smith SM, Flinn IW, Bartlett NL, et al. Magrolimab plus rituximab in relapsed/refractory indolent non-Hodgkin lymphoma: 3-year follow-up of a phase 1/2 trial. Blood Adv. 2024 Sep 3;8(22):5855-63. doi: 10.1182/bloodadvances.2024013277.
Link BK, Maurer MJ, Nowakowski GS, Ansell SM, Macon WR, Syrbu SI, et al. Rates and outcomes of follicular lymphoma transformation in the immunochemotherapy era: a report from the University of Iowa/MayoClinic Specialized Program of Research Excellence Molecular Epidemiology Resource. J Clin Oncol. 2013 Sep 10;31(26):3272-8. doi: 10.1200/JCO.2012.48.3990.
Wagner-Johnston ND, Link BK, Byrtek M, Dawson KL, Hainsworth J, Flowers CR, et al. Outcomes of transformed follicular lymphoma in the modern era: a report from the National LymphoCare Study (NLCS). Blood. 2015 Aug 13;126(7):851-7. doi: 10.1182/blood-2015-01-621375.
Marcus R, Imrie K, Solal-Celigny P, Catalano JV, Dmoszynska A, Raposo JC, et al. Phase III study of R-CVP compared with cyclophosphamide, vincristine, and prednisone alone in patients with previously untreated advanced follicular lymphoma. J Clin Oncol. 2008 Oct 1;26(28):4579-86. doi: 10.1200/JCO.2007.13.5376.
Jurinovic V, Passerini V, Oestergaard MZ, Knapp A, Mundt K, Araf S, et al. Evaluation of the m7-FLIPI in patients with follicular lymphoma treated within the Gallium trial: EZH2 mutation status may be a predictive marker for differential efficacy of chemotherapy. Blood. 2019 Nov 13;134(Suppl 1):122. https://doi.org/10.1182/blood-2019-130208
Cleary ML, Smith SD, Sklar J. Cloning and structural analysis of cDNAs for bcl-2 and a hybrid bcl-2/immunoglobulin transcript resulting from the t(14;18) translocation. Cell. 1986 Oct 10;47(1):19-28. doi: 10.1016/0092-8674(86)90362-4.
Tsujimoto Y, Cossman J, Jaffe E, Croce CM. Involvement of the bcl-2 gene in human follicular lymphoma. Science. 1985 Jun 21;228(4706):1440-3. doi: 10.1126/science.3874430.
Yunis JJ, Oken MM, Kaplan ME, Ensrud KM, Howe RR, Theologides A. Distinctive chromosomal abnormalities in histologic subtypes of non-Hodgkin’s lymphoma. N Engl J Med. 1982 Nov 11;307(20):1231-6. doi: 10.1056/NEJM198211113072002.
Graninger WB, Seto M, Boutain B, Goldman P, Korsmeyer SJ. Expression of Bcl-2 and Bcl-2-Ig fusion transcripts in normal and neoplastic cells. J Clin Invest. 1987 Nov 1;80(5):1512-5. doi: 10.1172/JCI113235.
Vogler M, Braun Y, Smith VM, Westhoff MA, Pereira RS, Pieper NM, et al. The BCL2 family: from apoptosis mechanisms to new advances in targeted therapy. Signal Transduct Target Ther. 2025 Mar 21;10(1):91. doi: 10.1038/s41392-025-02176-0.
Pasqualucci L, Bhagat G, Jankovic M, Compagno M, Smith P, Muramatsu M, et al. AID is required for germinal center-derived lymphomagenesis. Nat Genet. 2008 Jan;40(1):108-12. doi: 10.1038/ng.2007.35.
Correia C, Schneider PA, Dai H, Dogan A, Maurer MJ, Church AK, et al. BCL2 mutations are associated with increased risk of transformation and shortened survival in follicular lymphoma. Blood. 2015 Jan 22;125(4):658-67. doi: 10.1182/blood-2014-04-571786.
Rohlin A, Wernersson J, Engwall Y, Wiklund L, Björk J, Nordling M. Parallel sequencing used in detection of mosaic mutations: comparison with four diagnostic DNA screening techniques. Hum Mutat. 2009;30(6):1012-20. doi: 10.1002/humu.20980.
Correia C, Maurer MJ, McDonough SJ, Schneider PA, Ross PE, Novak AJ, et al. Relationship between BCL2 mutations and follicular lymphoma outcome in the chemoimmunotherapy era. Blood Cancer J. 2023 May 17;13(1):81. doi: 10.1038/s41408-023-00847-1.
Limpens J, Stad R, Vos C, de Vlaam C, de Jong D, van Ommen GJB, et al. Lymphoma-associated translocation t(14;18) in blood B cells of normal individuals. Blood. 1995 May 1;85(9):2528-36.
Morin RD, Johnson NA, Severson TM, Mungall AJ, An J, Goya R, et al. Somatic mutation of EZH2 (Y641) in follicular and diffuse large B-cell lymphomas of germinal center origin. Nat Genet. 2010 Feb;42(2):181-5. doi: 10.1038/ng.518.
Shilatifard A. Molecular implementation and physiological roles for histone H3 lysine 4 (H3K4) methylation. Curr Opin Cell Biol. 2008 Jun 1;20(3):341-8. doi: 10.1016/j.ceb.2008.03.019.
Morin RD, Mendez-Lago M, Mungall AJ, Goya R, Mungall KL, Corbett RD, et al. Frequent mutation of histone-modifying genes in non-Hodgkin lymphoma. Nature. 2011 Aug;476(7360):298-303. doi: 10.1038/nature10351.
Pasqualucci L, Dominguez-Sola D, Chiarenza A, Fabbri G, Grunn A, Trifonov V, et al. Inactivating mutations of acetyltransferase genes in B-cell lymphoma. Nature. 2011 Mar 10;471(7337):189-95. doi: 10.1038/nature09730.
Okosun J, Bödör C, Wang J, Araf S, Yang CY, Pan C, et al. Integrated genomic analysis identifies recurrent mutations and evolution patterns driving the initiation and progression of follicular lymphoma. Nat Genet. 2014 Feb;46(2):176-81. doi: 10.1038/ng.2856.
Hilton LK, Ngu HS, Collinge B, Dreval K, Ben-Neriah S, Rushton CK, et al. Relapse timing is associated with distinct evolutionary dynamics in diffuse large B-cell lymphoma. J Clin Oncol. 2023 Sep 1;41(25):4164-77. doi: 10.1200/JCO.23.00570.
Pasqualucci L, Khiabanian H, Fangazio M, Vasishtha M, Messina M, Holmes AB, et al. Genetics of follicular lymphoma transformation. Cell Rep. 2014 Jan 16;6(1):130-40. doi: 10.1016/j.celrep.2013.12.027.
Bouska A, Zhang W, Gong Q, Iqbal J, Scuto A, Vose J, et al. Combined copy number and mutation analysis identifies oncogenic pathways associated with transformation of follicular lymphoma. Leukemia. 2017 Jan;31(1):83-91. doi: 10.1038/leu.2016.175.
Kridel R, Chan FC, Mottok A, Boyle M, Farinha P, Tan K, et al. Histological transformation and progression in follicular lymphoma: a clonal evolution study. PLoS Med. 2016 Dec 13;13(12):e1002197. doi: 10.1371/journal.pmed.1002197.
Wright GW, Huang DW, Phelan JD, Coulibaly ZA, Roulland S, Young RM, et al. A probabilistic classification tool for genetic subtypes of diffuse large B cell lymphoma with therapeutic implications. Cancer Cell. 2020 Apr;37(4):551-568.e14. doi: 10.1016/j.ccell.2020.03.015.
Chakraborty S, Martines C, Porro F, Fortunati I, Bonato A, Marija Dimishkovska, et al. B-cell receptor signaling and genetic lesions in TP53 and CDKN2A/CDKN2B cooperate in Richter transformation. Blood. 2021 Sep 23;138(12):1053-66. doi: 10.1182/blood.2020008276.
Parry EM, Leshchiner I, Guièze R, Johnson C, Tausch E, Parikh SA, et al. Evolutionary history of transformation from chronic lymphocytic leukemia to Richter syndrome. Nat Med. 2023 Jan;29(1):158-69. doi: 10.1038/s41591-022-02113-6.
Martines C, Chakraborty S, Felician G, Hofmann K, Blasutig S, Negara I, et al. Richter syndrome-associated genetic lesions in TP53 and the cell cycle inhibitors CDKN2A and CDKN2B induce transformation of a CD5+ B cell subset characterized by a restricted IGHV repertoire, autoreactivity and BCR dependence. Blood. 2024 Nov 5;144(Suppl 1):759. https://doi.org/10.1182/blood-2024-207095
Chapuy B, Stewart C, Dunford AJ, Kim J, Kamburov A, Redd RA, et al. Molecular subtypes of diffuse large B cell lymphoma are associated with distinct pathogenic mechanisms and outcomes. Nat Med. 2018 May;24(5):679-90. doi: 10.1038/s41591-018-0016-8.
Chapuy B, Wood T, Stewart C, Dunford A, Wienand K, Khan SJ, et al. DLBclass: a probabilistic molecular classifier to guide clinical investigation and practice in diffuse large B-cell lymphoma. Blood. 2025 May 1;145(18):2041-2055. doi: 10.1182/blood.2024025652.
Stokes ME, Wenzl K, Huang CC, Ortiz M, Hsu CC, Maurer MJ, et al. Transcriptomic classification of diffuse large B-cell lymphoma identifies a high-risk activated B-cell-like subpopulation with targetable MYC dysregulation. Nat Commun. 2024 Aug 8;15(1):6790. doi: 10.1038/s41467-024-50830-y.
Negara I, Tomuleasa C, Buruiana S, Efremov DG. Molecular subtypes and the role of TP53 in diffuse large B-cell lymphoma and Richter syndrome. Cancers (Basel). 2024 Jan;16(12):2170. doi: 10.3390/cancers16122170.
Sarkozy C, Wu S, Takata K, Aoki T, Neriah SB, Milne K, et al. Integrated single cell analysis reveals co-evolution of malignant B cells and tumor micro-environment in transformed follicular lymphoma. Cancer Cell. 2024 Jun 10;42(6):1003-1017.e6. doi: 10.1016/j.ccell.2024.05.011.
Dreval K, Hilton LK, Cruz M, Shaalan H, Ben-Neriah S, Boyle M, et al. Genetic subdivisions of follicular lymphoma defined by distinct coding and noncoding mutation patterns. Blood. 2023 Aug 10;142(6):561-73. doi: 10.1182/blood.2022018719.
Pastore A, Jurinovic V, Kridel R, Hoster E, Staiger AM, Szczepanowski M, et al. Integration of gene mutations in risk prognostication for patients receiving first-line immunochemotherapy for follicular lymphoma: a retrospective analysis of a prospective clinical trial and validation in a population-based registry. Lancet Oncol. 2015 Sep;16(9):1111-22. doi: 10.1016/S1470-2045(15)00169-2.
Russler-Germain DA, Krysiak K, Ramirez C, Mosior M, Watkins MP, Gomez F, et al. Mutations associated with progression in follicular lymphoma predict inferior outcomes at diagnosis: Alliance A151303. Blood Adv. 2023 Sep 26;7(18):5524-39. doi: 10.1182/bloodadvances.2023010779.
Wu W, Bruscaggin A, Valera A, Flospergher E, Bonfiglio F, Spina V, et al. Evaluation of the different stratification models for POD24 prediction in patients with follicular lymphoma. Blood. 2020 Nov 5;136(Suppl 1):24-5. https://doi.org/10.1182/blood-2020-142000
Lockmer S, Ren W, Brodtkorb M, Østenstad B, Wahlin BE, Pan-Hammarström Q, et al. M7-FLIPI is not prognostic in follicular lymphoma patients with first-line rituximab chemo-free therapy. Br J Haematol. 2020;188(2):259-67. doi: 10.1111/bjh.16159.
Rodríguez-Sevilla JJ, Fernández-Rodríguez C, Bento L, Diez-Feijóo R, Pinzón S, Gibert J, et al. Evaluation of 4 prognostic indices in follicular lymphoma treated in first line with immunochemotherapy. Blood Adv. 2023 Apr 25;7(8):1606-14. doi: 10.1182/bloodadvances.2022007949.
Wright GW, Ramsower CA, Cerhan JR, Novak AJ, Link BK, Maurer MJ, et al. Prediction of early disease progression at 24 months (POD24) using pre-treatment biopsies from patients with follicular lymphoma (FL) treated with immunochemotherapy (IC). Blood. 2022 Nov 15;140(Supp 1):6426-7. https://doi.org/10.1182/blood-2022-160351
Freeman CL, Kridel R, Moccia AA, Savage KJ, Villa DR, Scott DW, et al. Early progression after bendamustine-rituximab is associated with high risk of transformation in advanced stage follicular lymphoma. Blood. 2019 Aug 29;134(9):761-4. doi: 10.1182/blood.2019000258.
Li X, Wang CY. From bulk, single-cell to spatial RNA sequencing. Int J Oral Sci. 2021 Nov 15;13(1):36. doi: 10.1038/s41368-021-00146-0.
Haebe S, Shree T, Sathe A, Day G, Czerwinski DK, Grimes SM, et al. Single-cell analysis can define distinct evolution of tumor sites in follicular lymphoma. Blood. 2021 May 27;137(21):2869-80. doi: 10.1182/blood.2020009855.
Araf S, Wang J, Korfi K, Pangault C, Kotsiou E, Rio-Machin A, et al. Genomic profiling reveals spatial intra-tumor heterogeneity in follicular lymphoma. Leukemia. 2018;32(5):1261-5. doi: 10.1038/s41375-018-0043-y.
Scherer F, Kurtz DM, Newman AM, Stehr H, Craig AFM, Esfahani MS, et al. Distinct biological subtypes and patterns of genome evolution in lymphoma revealed by circulating tumor DNA. Sci Transl Med. 2016 Nov 9;8(364):364ra155. doi: 10.1126/scitranslmed.aai8545.
Andor N, Simonds EF, Czerwinski DK, Chen J, Grimes SM, Wood-Bouwens C, et al. Single-cell RNA-Seq of follicular lymphoma reveals malignant B-cell types and coexpression of T-cell immune checkpoints. Blood. 2019 Mar 7;133(10):1119-29. doi: 10.1182/blood-2018-08-862292.
Attaf N, Cervera-Marzal I, Gil L, Dong C, Navarro JM, Spinelli L, et al. Single-cell RNA sequencing identifies a pseudo-immune differentiation axis as the main source of functional heterogeneity in follicular lymphoma B-cells. Blood. 2019 Nov 13;134(Suppl 1):548. https://doi.org/10.1182/blood-2019-125718
Radtke AJ, Roschewski M. The follicular lymphoma tumor microenvironment at single-cell and spatial resolution. Blood. 2024 Mar 21;143(12):1069-79. doi: 10.1182/blood.2023020999.
Dave SS, Wright G, Tan B, Rosenwald A, Gascoyne RD, Chan WC, et al. Prediction of survival in follicular lymphoma based on molecular features of tumor-infiltrating immune cells. N Engl J Med. 2004 Nov 18;351(21):2159-69. doi: 10.1056/NEJMoa041869.
Han G, Deng Q, Marques-Piubelli ML, Dai E, Dang M, Ma MCJ, et al. Follicular lymphoma microenvironment characteristics associated with tumor cell mutations and MHC class II expression. Blood Cancer Discov. 2022 Sep 6;3(5):428-43. doi: 10.1158/2643-3230.BCD-21-0075.
Radtke AJ, Kandov E, Lowekamp B, Speranza E, Chu CJ, Gola A, et al. IBEX: A versatile multiplex optical imaging approach for deep phenotyping and spatial analysis of cells in complex tissues. Proc Natl Acad Sci USA. 2020 Dec 29;117(52):33455-65. doi: 10.1073/pnas.2018488117.
Radtke AJ, Postovalova E, Varlamova A, Bagaev A, Sorokina M, Kudryashova O, et al. Multi-omic profiling of follicular lymphoma reveals changes in tissue architecture and enhanced stromal remodeling in high-risk patients. Cancer Cell. 2024 Mar 11;42(3):444-463.e10. doi: 10.1016/j.ccell.2024.02.001.